УНИКАЛЬНЫЙ РАЗРЕЗ БЕЛГОРОДСКОГО ЧЕРНОЗЕМА
Комментарии фото почвенного разреза «Ямская степь» заповедника Белогорье.
А.С.Керженцев
Институт фундаментальных проблем биологии РАН, Пущино Московской обл.
Для участников полевой экскурсии VII съезда Общества почвоведов им. В.В. Докучаева и Всероссийской с зарубежным участием научной конференции «Почвоведение – продовольственной и экологической безопасности страны» Белгород, 15–22 августа 2016 г. в заповеднике Белогорье урочище «Ямскую степь» организаторы подготовили уникальный разрез сложноорганизованной почвенно-седиментационной толщи, сформированной в течение последнего макроцикла (~150 тыс. л.н.), то есть в позднем неоплейстоцене и голоцене. Вскрытая мощность разреза составляет ~6,5 м, ширина описанной южной стенки 4 м. Прекрасный обзор всех деталей обнажения.
Авторы диагностировали следующие дневные почвы, палеопочвы и педокомплексы, в различной степени подверженные позднеплейстоценовым криогенным процессам в виде клиньев и трещин:
- Голоценовый целинный миграционно-мицелярный чернозем на поздневалдайских
(МИС2) карбонатных лессовидных суглинках (Морская изотопная стадия (МИС) МИС1);
- Два ритма брянской палеопочвы (МИС3), совмещенные с нижней частью
профиля дневного чернозема;
- Ранневалдайские педоседименты инициального полугидроморфного педогенеза
(2 педоседимента) и подстилающая их супесчаная пачка (МИС4);
- Темногумусовая глеевая почва и перекрывающие её два глеевых горизонта
(МИС5а-d);
- Микулинская почва (МИС5e), представленная серией горизонтов [BTg1]-[BTg2]-[Gt];
- в основании разреза – вскрытое залегание олигоценовых песков. (Путеводитель, 2016).
Уникальность разреза (Фото-1) в фиксированной границе начала голоценовых отложений. Если бы этот разрез увидели Н.А.Богословский, Л.С.Берг, С.С.Неуструев, П.С.Коссович, Б.С.Личков, Р.С.Ильин они бы с полной уверенностью доказали выдвинутую ими гипотезу о почвенном происхождении лессов. В путеводителе, кроме фото профиля, имеется его рисунок с указанием уточненных художником размеров целого ряда горизонтов и их описанием вплоть до определения точного места в шкале геологического времени. Однако рисунок все-таки содержит субъективную оценку структуры горизонтов и частных деталей профиля конкретным человеком с его конкретным опытом и багажом знаний. Столь уникальный разрез должен иметь несколько альтернативных описаний, тогда его ценность для науки многократно возрастет.
Мы попытались заочно, на основе фотоснимков и описаний осуществить альтернативную интерпретацию описания этого разреза с позиции функциональной экологии – науки изучающей механизм функционирования экосистем и почв как их незаменимых компонентов (Ковда, 1968, 1974; Ковда и др., 1990; Керженцев, 2006, 2012); с позиции «почвы-момента» (Соколов, Таргульян, 1976), а говоря точнее, с позиции «физиологии» экосистем.
Конечно, по фотоснимку без прямого контакта с реальным объектом сложно определить состав и границы горизонтов, поэтому мы попытались использовать результаты уже имеющихся описаний с некоторым укрупнением выделенных авторами слоев. При необходимости это не трудно уточнить и поправить. Для нас было важно определить мощность горизонтов и вычислить время, в течение которого произошло накопление этой мощности.
Самый нижний слой разреза 600-650 см представлен песчаными отложениями. На них залегает палеопочва Микулинского межледниковья мощностью 180 см (420-600 см) с явными признаками криогенеза. Обилие темного органического вещества указывает на захоронение большой массы растительных остатков возможно торфяно-болотной почвы. Этот богатый органикой слой перекрывается светлой прослойкой совсем иного состава и неясного генезиса. Извилистая нижняя и ровная верхняя граница слоя может быть результатом осаждения взвесей мутных талых вод, образованных при таянии льда отступающего ледника с заполнением депрессий, сформированных при вытаивании жильного льда и с выравненной поверхностью –дном медленных водотоков. Поскольку на снимке эта часть плохо просматривается, мы ограничимся этими замечаниями и рассмотрим подробнее вышележащую толщу.
Вся вышележащая толща 420 см сформировалась в течение периода голоцена, который продолжается 10(12) тыс. лет, после отступления последнего поздневалдайского или Осташковского оледенения с его холодным фронтом климатических условий, который простирался на тысячу и более километров к югу от края ледника (Величко, 1972).
По мере отступления ледника вслед за ним с юга на север стали перемещаться климатические и природные зоны (Берг, 1950). Последовательность почв в разрещзе, сформированная в ходе осадконакопления показывает, что сначала на месте «Ямской степи» сформировались таежные экосистемы с подзолистыми почвами, потом их сменили хвойно-широколиственные леса с дерново-подзолистыми почвами, затем здесь сформировались лесостепи с серыми лесными почвами, которые сменились мезофильными лугами и луговыми степями с темно-серыми лесными почвами и выщелоченными черноземами. И, наконец, на данной территории обосновались настоящие степи с типичными черноземами, которые существуют здесь до сих пор.
Это соответствует гипотезе Б.Л.Личкова (1945), который подтвердил высказанные ранее суждения Н.А.Богословского (1899), Л.С.Берга (1916), С.С.Неуструева (1926), П.С.Коссовича (1911), А.Н.Соколовского (1933) о почвенном происхождении лесса, каждый тип почвы порождает только ему соответствующий тип подстилающих пород: под красноземом образуются латериты, под лесными почвами – покровные суглинки, под черноземами – лессы, под каштановыми почвами – шоколадные глины. Этим они объясняли отмеченную многими геологами и почвоведами зональность материнских почвообразующих пород, которые на самом деле являются дочерними почвообразованными породами. Состав седиментов зависит от типа почв и экосистем, а мощность слоев – от времени существования на данной территории экосистем и почв данного типа (Личков, 1945). Следовательно все горизонты, вскрытые данным разрезом сформировались соответствующими типами почв, которые сменяли друг друга по мере отступления ледника и потеплении климата.
Если за 10 тысяч лет, прошедших от начала периода голоцена до наших дней на данной территории накопилась толща седиментов 420 см, значит, средняя скорость седиментации была 0,04 см/год или 40 см за 1000 лет. Отсюда следует, что при постоянной во времени скорости поступления осадочного материала накопление отдельных слоев происходило снизу вверх, по мере отложения почвами отходов метаболизма экосистемы, в течение следующих отрезков времени:
Вся толща 0-420 см мощностью 420 см накопилась в течение 10500 лет.
Слой 420-370 см (50 см) накопился за 1250 лет, с 10500 по 9250 лет назад.
Слой 370-320 см (50 см) накопился за 1250 лет с 9250 по 8000 лет назад..
Слой 320-260 см (60 см) накопился за 1500 лет с 8000 по 6500 лет назад..
Слой 260-220 см (40 см) накопился за 1000 лет с 6500 по 5500 лет назад.
Слой 220-0 см (220 см) накопился в течение 5500 лет с 5500 до наших дней.
Нижний горизонт голоцена 420-370 см начал формироваться 10500 лет назад в результате метаболизма экосистем тундры и лесотундры и закончил свое накопление через 1250 лет то есть 9250 лет назад. Выше, на глубине 370-320 см залегает горизонт, который формировался в диапазоне 1250-8000 лет подзолистыми почвами под хвойными лесами в условиях климата более теплого по сравнению со временем отложения подстилающего слоя. По мере дальнейшего потепления климата сформировались хвойно-широколиственные леса с дерново-подзолистыми почвами. Эти экосистемы отложили горизонт 370-320 см, который осаждался в течение 1250 лет и сменился 8000 лет назад следующим горизонтом 320-260 см, который отложили экосистемы широколиственных лесов с серыми лесными почвами в течение 1500 лет от 8000 до 6500 лет назад. Следующий горизонт 260-220 см сформировался за 1000 лет в результате метаболизма лесостепных экосистем с темно-серыми лесными почвами и выщелоченными черноземами, которые откладывали свои отходы – седименты от 6500 до 5500 лет назад.
После этого на территории «Ямской степи» установились стабильные климатические условия климатического оптимума с устойчивой природной зональностью, освободившейся от влияния последнего Осташковского оледенения. В этих условиях сформировались настоящие степи с типичными черноземами, которые в течение 5500 лет до настоящего времени отложили толщу лессовидных суглинков мощностью 220 см. Однако первые 500 лет потребовались для накопления метровой толщи гумусового профиля чернозема, который сохранился до сих пор благодаря периодическому обновлению почвенного органического вещества за счет ежегодного поступления растительного и животного опада в почву и последовательной минерализации.
Накопление толщи происходило за счет увеличения мощности горизонта С, который и сейчас продолжает нарастать ежегодно со скоростью 0,04 см/год, то есть 40 см за 1000 лет. При этом мощность верхних гумусовых горизонтов чернозема (горизонты А и В) сохраняется стабильно, поскольку при стабильном климате каждые 350 лет происходит полное обновление его органического профиля в стационарном режиме (Ковда и др., 1990). За это время все фракции гумуса последовательно сменяются новыми фракциями в результате ежегодного поступления в почву и переработки педоценозом свежего опада отмершей биомассы, последующего синтеза и минерализации фракций гумуса. Голоцен – считается периодом климатического оптимума, который находится между крайностями периодов оледенения и аридизации. Колебания климата в пределах оптимального диапазона сопровождаются флуктуациями параметров экосистем, то есть их количественными изменениями в пределах диагностики таксона.
Вышеизложенное позволяет высказать сомнение по поводу радиоуглеродной датировки верхних горизонтов разреза. Поскольку органическое вещество в метаболизме экосистемы многократно переходит из одного состояния в другое, измерение возраста почвы радиоуглеродным методом нельзя признать корректным. По нашим расчетам (Ковда и др., 1990) гумус подзолистых почв обновляется каждые 70-80 лет, серых лесных – 100-120 лет, черноземов 350-500 лет, красноземов – 10-20 лет. При такой динамичности измеряемый углерод мог много раз побывать в составе почвы, растений, животных, перемещаясь по трофическим цепям, циркулируя в биологическом круговороте. Он не мог лежать на месте в течение тысяч лет. В живой почве (педоценозе), которая является непременным и незаменимым и постоянно действующим компонентом экосистем (Ковда, 1968, 1974), все элементы постоянно перемещаются в цикле метаболизма. Этот круговорот сопровождается разделение изотопов. Как и для других элементов, изотопное разделение углерода в биологическом круговороте может приводить к существенным ошибкам датирования. Поэтому приведенные данные о возрасте горизонтов чернозема 25-28 тыс. лет вызывают сомнение. В живой почве даже самая устойчивая фракция гумуса – гумины, обновляется каждые 350-500 лет (Ковда и др., 1990).
Более точное и тщательное описание горизонтов на стенке разреза может изменить время их накопления, но не принципиально. Важно то, что в каждый период времени данная территория находилась под влиянием конкретного диапазона климатических условий, к которому адаптировались конкретные типы экосистем с соответствующими типами почв. Каждый тип экосистем и почв откладывал отходы своего метаболизма в форме глинистых кутан, железо-марганцевых и карбонатных конкреций, вторичных и первичных минералов определенного состава. Отходы метаболизма экосистем необратимо переходили из биологического круговорота в геологический, образуя слои осадочных пород.
Б.Л.Личков (1945, С.548). «Внимание исследователей – геологов и почвоведов, - когда они касались связи почвы и горных пород, привлекал в основном вопрос о том, как разлагаются и разрушаются материнские горные породы, производя почву; иными словами, это был вопрос о том, как материнские горные породы создают почвенный покров. Хотя последний процесс существует, но существует он в особых условиях, условиях денудации; фактически же та порода, на которой непосредственно лежит почва, вовсе никогда не разрушалась, производя почву, ибо отношение между ними как раз обратное: не порода произвела почву, а почва систематически, постепенно создавала ту породу, которая ее подстилает… по мере нарастания толщи этих пород почвенный покров неизбежно должен подниматься все выше и выше, лежа все время над слоем породы, но это отнюдь не значит, что он этой породой впервые выделен. Скорее, наоборот, можно сказать, что в начале существования данного почвенного покрова подлежащей ему породы еще не было, между тем почвенный покров существовал, являясь по составу своему вначале продуктом выветривания предыдущей породы».
Каждый тип экосистем формировал свой тип седиментов определенного механического и минералогического состава: гилеи с красноземами откладывали латериты, лесные экосистемы умеренного пояса с подзолистыми, бурыми и серыми почвами – покровные суглинки, степные экосистемы с черноземами – лессы, полупустыни с каштановыми почвами – шоколадные глины. Зональные почвы сформировали зональность материнских «почвообразующих» пород, которая была отмечена многими почвоведами и геологами еще в начале прошлого века (Л.С.Берг, С.С.Неуструев, П.С.Коссович, А.Е.Ферсман, Б.Б.Полынов и другие). На самом деле осадочные породы являются дочерними почвообразованными породами (Личков, 1945). Данный разрез служит прямым доказательством справедливости гипотезы Б.Л.Личкова (1945, С.551): «Каждая почва всегда творила и творит непосредственно под собой новые породы, производя отложение осадков из своих почвенных растворов; именно поэтому под каждой зональной разновидностью почв породы, естественно, должны быть иными, откуда вытекает, что творимый почвами осадочный покров пород, очевидно, тоже должен носить неизбежно черты зональности». Ниже на рис.1 показан механизм образования седиментов в процессе метаболизма экосистем.
Механизм образования осадочных пород как отходов метаболизма экосистем удалось описать на основе новых знаний, полученных в рамках функциональной экологии – науки, изучающей механизм функционирования экосистем, незаменимым компонентом которых является почва, а точнее педоценоз (В.А.Ковда, 1968, 1974, 1990; Керженцев, 2006, 2012, 2014). Экосистема –симбиотическая ассоциация трех биоценозов, специализированных по типам питания: фитоценоза, зооценоза и педоценоза. Эта ассоциация функционирует автономно за счет обмена симбионтов отходами жизнедеятельности. Кооперация биоценозов избавила их от энергетических затрат на поиск и добывание пищевых ресурсов. Каждый компонент экосистемы получает их от партнеров в форме отходов в обмен на свои отходы. Фитоценоз превращает минеральные элементы в живую фитомассу, зооценоз превращает часть фитомассы в зоомассу, После отмирания растений и животных их масса превращается в мертвую некромассу, которую сапротрофная биота педоценоза превращает в минеральные элементы, необходимые фитоценозу для фотосинтеза. Цикл метаболизма экосистемы превратил ограниченный запас биофильных элементов в постоянно обновляющийся, а потому бесконечный ресурс. Степень замкнутости цикла метаболизма современных экосистем 90-99% их общей массы (Марчук, Кондратьев, 1992; Горшков, 1995). Это значит, что отходы метаболизма экосистем составляют 1-10% их экомассы. Для поддержания геохимического баланса эти потери должны компенсироваться притоком вещества извне. Эта компенсация осуществляется за счет атмосферных, в том числе метеоритных выпадений и продуктов выветривания горных пород. Состав метеоритного вещества и космической пыли, которая осаждается на земную поверхность со средней скоростью 0,6 т/км2/год в течение 4,5 млрд лет, очень близок составу биомассы и далек от состава геологических пород (Голенецкий и др., 1981).
Отсутствие полной замкнутости цикла метаболизма объясняется тем, что порядок потребления элементов при синтезе фитомассы не совпадает с порядком их высвобождения при минерализации некромассы. Поэтому всегда остаются выделенные педоценозом, но невостребованные фитоценозом элементы, которые при накоплении могут оказать токсичное воздействие на биоту экосистемы. Этого не происходит потому, что свободные элементы взаимодействуют с органическими радикалами разлагающейся некромассы и образуют гумусовые соединения, которые связывают потенциальную токсичность свободных элементов и хранят их до востребования фитоценозом.
Растения, испытывающие дефицит минерального питания с помощью корневых выделений провоцируют вспышку численности прикорневой микрофлоры, которая быстро «съедает» приманку и переключается на потребление гумуса, а минеральные элементы, выделяемые микрофлорой в качестве отходов, компенсируют дефицит питания растений. При этом растения усваивают только часть элементов, высвобождаемых при минерализации гумуса. Не усвоенные фитоценозом элементы, способные при накоплении оказать токсическое воздействие на биоту, подвергаются биокристаллизации и в форме глинистых кутан, железо-марганцевых и карбонатных конкреций, вторичных и первичных минералов выпадают из биологического круговорота. «Лишние» элементы, связанные прочно кристаллической решеткой, представляют абсолютные отходы метаболизма экосистемы, которые необратимо переходят в геологический круговорот и формируют минеральные седименты. Сначала они разбавляют органическую массу опада и гумуса, затем формируют минеральную основу почвенного профиля, а со временем постепенно накапливаются в виде подпочвенного горизонта С и слоя осадочных пород.
А начинается все с растительного и животного опада отмершей биомассы, которую почвенная биота превращает в минеральные элементы, необходимые растениям для фотосинтеза. Животные превращают часть фитомассы в зоомассу, которую после отмирания почвенная биота также как фитомассу превращает в минеральную массу – пищу фитоценоза. Продуктами минерализации отмершей биомассы являются газы, растворы и коллоиды. Основную их массу поглощает фитоценоз, а не усвоенные газы пополняют состав атмосферы, растворы – состав гидросферы, коллоиды состав литосферы.
Рисунок 1 иллюстрирует механизм нейтрализации «излишка» элементов в процессе метаболизма экосистем путем их гумификации и биокристаллизации. Живая биомасса по завершении жизненного цикла превращается в некромассу (опад) и репродуктивную массу (плоды, семена, споры, зародыши). Репродуктивная масса превращается в новую биомассу в следующем поколении, а растительный и животный опад разрушается сапротрофной биотой с выделением газов, растворов и органических радикалов. Газы поглощаются листвой растений, растворы – корнями, а свободные органические радикалы связывают неусвоенные фитоценозом элементы, образуя фульваты – первые гумусовые фракции. Устойчивая к разложению часть опада накапливается в форме лесной подстилки или степного войлока. Эту полуразложившуюся массу использует в пищу почвенная мезофауна, способная разрушить устойчивые фракции некромассы. Часть выделенных при этом минеральных элементов усваивается фитоценозом, а часть взаимодействует с органическими радикалами и синтезирует новые органо-минеральные соединения в форме гуматов, составляющих основу почвенного гумуса. Гумификация защищает биоту экосистемы от потенциальной токсичности свободных элементов и хранит эти элементы до востребования фитоценозом. Гумусовые фракции в свою очередь становятся пищей для почвенной микрофлоры, которая высвобождает в качестве отходов связанные в них минеральные элементы. Часть элементов усваивает фитоценоз, а неусвоенная часть подвергается биокристаллизации и в форме глинистых кутан, железо-марганцевых и карбонатных конкреций, вторичных и первичных минералов формирует минеральную основу почвенного профиля, а со временем накапливает подпочвенный горизонт С и слой осадочных пород.
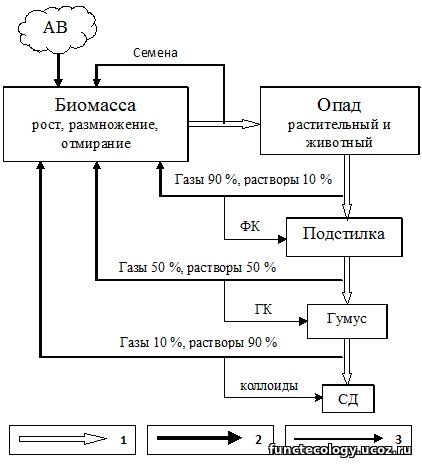
Рис. 1. Утилизация отходов метаболизма экосистемы.
1) стадии превращения отмершей биомассы; 2) поступление вещества в биомассу; 3) утилизация химических элементов, невостребованных фитоценозом.
Глубокое проникновение карбонатных и железо-марганцевых конкреций, заметные в нижней части разреза на глубине 420-370 см объясняется наличием в подпочвенных отложениях водопроводящих каналов и пустот (трещин, ходов корней, насекомых и землероев). При минерализации биомассы высвобождаются легко растворимые бикарбонаты, закиси железа и марганца, которые вместе с почвенной влагой пропитывают минеральную толщу на определенную глубину. В процессе высыхания профиля растворы мигрируют к испаряющим поверхностям, не обязательно к дневной поверхности. В глубоких горизонтах таковыми являются трещины усыхания, морозобойные трещины, ходы корней, насекомых и землероев, которые заполняются воздухом, при контакте с ним бикарбонаты становятся нерастворимыми карбонатами, а закиси железа и марганца – нерастворимыми окислами, и выпадают в осадок. Бывшая сеть мелких корней образует карбонатный псевдомицелий, отмеченный в описании разреза, в ходах более крупных корней, мелких землероев, пустотах от куколок насекомых образуется белоглазка, журавчики, бобовины, куколки в зависимости от концентрации растворов, размеров и форм испарительных полостей. Многократное повторение миграционных процессов почвенной влаги накапливает массу новообразований в нижних горизонтах профиля в форме кутан, конкреций, примазок.
Почва как компонент экосистемы функционирует вместе с другими компонентами ритмично в суточном, годовом и многолетнем циклах. Она реагирует на все изменений экологических условий, но эта реакция не заметна на морфологическом профиле почвы, который поддерживается в стабильном состоянии благодаря постоянной работе механизма функционирования – метаболизма экосистемы в стационарном режиме. Этот механизм живо реагирует на внешние воздействия как естественных, так и антропогенных факторов. Причем естественные факторы (свет, тепло, влага, аэрация) оказывают преимущественное влияние на функцию метаболизма экосистемы и поддерживают ее устойчивость, а антропогенные факторы – на структуру экосистемы и стимулируют ее деградацию. Для изучения динамических процессов в почвах нужна принципиально новая экспериментальная база, способная контролировать изменчивость параметров педоценоза во времени. Она позволит перенять у природы безопасные технологии управления циклом метаболизма, который является универсальным механизмом функционирования всех живых систем: клетки, многоклеточного организма, экосистемы, биосферы. Сочетание «синтез-распад» безотказно действует на всех уровнях организации жизни. Почвоведы и геологи тщательно изучают процессы распада, игнорируя процессы синтеза которые строго согласованы друг с другом.
Б.Л.Личков (1945, С.548) справедливо сказал: «Внимание исследователей – геологов и почвоведов, - когда они касались связи почвы и горных пород, привлекал в основном вопрос о том, как разлагаются и разрушаются материнские горные породы, производя почву; иными словами, это был вопрос о том, как материнские горные породы создают почвенный покров. Хотя последний процесс существует, но существует он в особых условиях, условиях денудации; фактически же та порода, на которой непосредственно лежит почва, вовсе никогда не разрушалась, производя почву, ибо отношение между ними как раз обратное: не порода произвела почву, а почва систематически, постепенно создавала ту породу, которая ее подстилает… по мере нарастания толщи этих пород почвенный покров неизбежно должен подниматься все выше и выше, лежа все время над слоем породы, но это отнюдь не значит, что он этой породой впервые выделен. Скорее, наоборот, можно сказать, что в начале существования данного почвенного покрова подлежащей ему породы еще не было, между тем почвенный покров существовал, являясь по составу своему вначале продуктом выветривания предыдущей породы». В природе гармонично уравновешены процессы синтеза и распада. Но синтез признается для растений и животных, а почва считается разрушителем некромассы, хотя гумус, почвенные конкреции и вторичные минералы несомненно являются результатом биосинтеза.
После любезного предоставления О.С.Хохловой дополнительных фотоснимков с изображением нижней части разреза мы попытались внести коррективы в описание нижней толщи разреза мощностью 650-420 см.
Основание разреза 600-650 см представлено песчаными отложениями с округлыми светлыми пятнами размером 20-40 см. Этот слой (Фото-2) образовался в результате механического выветривания скальных пород под прямым воздействием ледника под его подошвой или в непосредственной близости от края ледника. Светлые округлые пятна – это валуны, образованные из обломков скальных пород на месте залегания под влиянием криогенеза. В почвенных разрезах восточного Забайкалья мы вскрывали валуны биотитового гранита разной размерности (от 10-20 до 40-60 см), которые сохраняли внутреннюю структуру и внешний облик, но легко разрезались ножом и лопатой. Криогенные условия (высокая влажность и резкие колебания температур) выпучивали на дневную поверхность засыпанные рыхлыми отложениями крупные обломки скальных пород, придавая им округлую форму. Часть валунов, которые остались на глубине после отступления контрастных климатических условий криогенеза потеряла внутренние связи минералов в структуре гранита. Валуны размером 40-60 см обнаруженные на глубине 100-150 см в почвенных разрезах черноземов в долине р. Шилка в районе г.Нерчинска, сохраняли форму и внутреннюю структуру валуна биотитового гранита, но легко разрезались ножом и лопатой. Разрушение внутренней связи минералов гранита происходило после отступления условий активного криогенеза. (Рис.2).
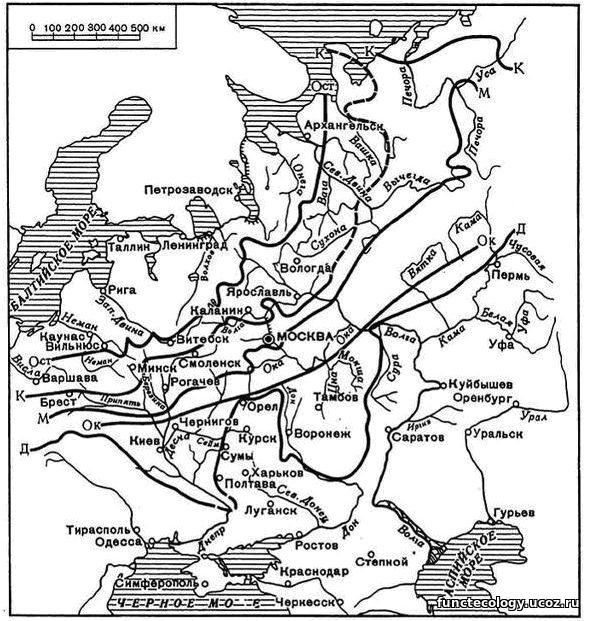
Рис.2. «Классические» границы распространения древних покровных оледенений на территории Восточно-Европейской платформы:
Ок–Ок – предполагаемая граница окского оледенения;
D–D – граница распространения днепровского оледенения;
М–М – граница распространения московского оледенения;
К–К – граница распространения валдайского оледенения;
Ост – граница распространения осташковского оледенения.
На территории «Ямской степи» активный криогенез мог быть в периоды Окского и Днепровского оледенений, когда край ледника находился в непосредственной близости. Постепенное отступление фронта Днепровского оледенения до границ Московского, Валдайского и Осташковского оледенений создавало благоприятные условия для перемещения природных зон с юга в высокие широты. Сначала здесь могли сформироваться экосистемы полярных пустынь, потом тундровые и лесотундровые экосистемы с мощными торфяниками, которые образовали гигантские залежи железных руд (КМА). При дальнейшем отступлении ледника здесь сформировались экосистемы хвойных, а затем хвойно-широколиственных лесов. Только после полного схода Осташковского оледенения и установления климатического оптимума здесь сформировались лесостепные, а затем степные экосистемы, которые господствуют на данной территории до настоящего времени и в течение 5-6 тысяч лет сформировали толщу лессовидных суглинков мощностью 420 см.
Слой 600-420 см сформировался в условиях активного криогенеза (Фото-3) . Об этом говорят округленные очертания полигональных блоков и заполненные органическим веществом полигональные трещины. Обилие погребенного органического вещества может быть результатом накопления торфа торфяно-болотными почвами. По данным болотоведов (Елина и др., 1984) накопление торфа происходит со скоростью 0,3 – 0,15 мм/год. В метаболизме болотных экосистем торф превращается в сапропель, а сапропель в глей – минеральную массу глинистого мехсостава (оглинение). Если принять минимальную скорость прироста торфа 0,15 мм/год, а его превращения в сапропель и глей 0,1 мм/год, то можно предположить, что слой 180 см (600-420 см) мог накопиться в течение 18000 лет. Началось его накопление 28500 лет назад и завершилось 10500 лет назад, когда начался период голоцена. Вполне возможно, что именно этот углерод, пройдя несколько циклов метаболизма разных типов экосистем, оказался в составе гумуса верхних горизонтов современного чернозема, возраст которых оказался равным по радиоуглеродной датировке 25-28 тыс. лет.
Самые нижние 50 см профиля (650-600 см) накапливались с гораздо меньшей скоростью в процессе механического выветривания скальных пород в суровых условиях арктического климата. Скорость накопления песчаного материала в результате механического разрушения обломков скальных пород на порядок меньше скорости накопления глея в болотных экосистемах, тоесть 0,01 мм/год. Поэтому можно считать, что вскрытая разрезом толща 50 см накапливалась в течение 50000 лет, а данный разрез демонстрирует процесс формирования толщи осадков на протяжении 50 тысяч лет. Приведенные расчеты не претендуют на точность датировок накопления слоев седиментов, для этого нужны более тщательные и точные измерения. Целью данного сообщения было показать работу механизма накопления седиментов осадочных пород как отходов метаболизма экосистем и подтвердить реальность гипотезы Б.Л.Личкова, Н.А.Богословского, Л.С.Берга, С.С.Неуструева, П.С.Коссовича о почвенном происхождении некоторых осадочных пород (лессов, покровных суглинков, латеритов) в плакорных условиях водораздельных ландшафтов, исключающих денудацию и переотложение осадков.
Геологи уже доказали, что строматолиты образовались в результате жизнедеятельности циано-бактериальных матов (Кузнецов, 2015). Осталось сделать следующий шаг и доказать, что современные экосистемы оставляют более существенный след в геологической истории Земли в виде характерных слоев осадочных пород. Морские осадки легко идентифицируются по захоронению раковин моллюсков. Время континентальных отложений определяется по отпечаткам листьев, насекомых, микроорганизмов, сохранившимся в метаморфических породах в разных регионах планеты на разной глубине от земной поверхности. В этих находках много случайностей, а по ним датируются целые эпохи. Гораздо проще было бы определить возраст слоев по их химическому, минералогическому и механическому составу. Но для этого необходимо более тщательно изучить образование современных седиментов типичными экосистемами мира. Надо узнать каким образом отмершая биомасса превращается в почвенный гумус горизонта А, который в результате минерализации превращается в горизонт В, а тот в результате биокристаллизации становится горизонтом С современных почв, то есть превращается в осадочную породу. Тогда можно будет читать в конкретном геологическом обнажении календарь природы, записанный в разноцветных слоях стратиграфической колонки. Состав и цвет слоя позволит определить тип экосистем, существовавших в данной местности в определенное время, а мощность слоя укажет на продолжительность периода, в течение которого этот тип экосистемы существовал в условиях конкретного типа климата и накопил данный слой седиментов. Геологические обнажения и разрезы станут более информативными, а информация более надежной.
Заключение
Классический пример формирования чернозема на стенах Староладожской крепости приведен самим В.В.Докучаевым. Но главное внимание уделено мощности гумусового горизонта, а его минеральная основа не рассматривалась. Практика почвоведения уже накопила множество примеров формирования современных почв на отвалах горных пород, сформировало новое направление – археологическое почвоведение, изучающее погребенные почвы. Однако представление и почве как геологической породе, преобразованной факторами почвообразования оказалось сильнее новой парадигмы – почва –незаменимый компонент биосферы и каждой ее экосистемы, которую выдвинул и обосновал еще в 1968 году В.А.Ковда (1971). Поэтому чиновники легко отражают попытки почвоведов пробить Закон об охране почв от их отчуждения, загрязнения и деградации. Самые продвинутые из них заявляют: мы знаем, что почва – это геологическая порода, преобразованная пятью факторами почвообразования. Мы видим, что геологических пород у нас вполне достаточно, а все пять факторов имеются в наличии. Мы не видим причин для беспокойства, нам достаточно Земельного кодекса.
Но почва это живое вещество – педоценоз, биологический реактор, который перерабатывает отмершую биомассу в минеральные элементы, необходимые фитоценозу. Тройственный симбиоз фитоценоза, зооценоза и педоценоза сформировал экосистему, функционирующую автономно за счет обмена симбионтов отходами жизнедеятельности. Метаболизм экосистемы избавил участников от энергетических затрат на поиск и добывание пищевых ресурсов, каждый получает их от партнеров в форме отходов в обмен на собственные отходы.
Метаболизм экосистемы не только обеспечивает компонентов пищевыми ресурсами, он решает еще одну жизненно важную функцию – утилизацию отходов жизнедеятельности. Поскольку порядок потребления элементов в процессе фотосинтеза не совпадает с порядком их высвобождения при деструкции некромассы, образуются «лишние» элементы, способные при накоплении оказать токсическое воздействие на биоту экосистемы. Свободные элементы взаимодействуют с органическими радикалами разлагающейся некромассы, образуя устойчивые органо-минеральные соединения с общим названием – гумус, который хранит химически связанные элементы до их востребования фитоценозом. Растения, испытывающие дефицит минерального питания с помощью корневых выделений провоцируют вспышку численности прикорневой микрофлоры, которая быстро «съедает» приманку и переключается на гумус, высвобождая из него минеральные элементы. Растения поглощают только нужную им часть освобожденных из гумуса элементов. Невостребованные фитоценозом и потенциально токсичные элементы подвергаются биокристаллизации и в форме глинистых кутан, железо-марганцевых и карбонатных конкреций, вторичных и первичных минералов пополняют минеральную основу почвенного профиля, которая со временем накапливает слои осадочных пород. По мере накопления осадко, они погружаются вглубь земной коры и под влиянием высоких давлений и температур превращаются в метаморфические породы (печаники, сланцы, алевролиты и др. Дальнейшее погружение их в мантию Земли переплавляет их в магматические породы, которые выносятся тектоническими процессами на земную поверхность в виде горных хребтов. Этим завершается цикл геологического круговорота.
Литература
- Берг Л.С. О происхождении лесса. Изв. РГО, т.52, вып.8, 1916. с.579-646.
- Берг Л.С. Некоторые соображения о послеледниковых изменениях климата и о лесостепье. Вопр. Геогр. 1950. сб.23. с. 57-84.
- Богословский Н.А. О некоторых явлениях выветривания в области Русской равнины. Изв. Геол. ком-та. Т.18. №5. СПб.1899.
- Горшков В.Г. Физические и биологические основы устойчивости жизни. М.: ВИНИТИ, 1995. 470 с.
- Елина Г.А., Кузнецов О.А., Максимов А.И. Структурно-функциональная организация и динамика болотных экосистем Карелии. Л.:Наука, 1984.
- Симакова А.Н., Пузаченко А.Ю. Реконструкция растительного покрова Русской равнины второй половины позднего неоплейстоцена и среднего голоцена. С 404-430. Биосфера-экосистема-биота в прошлом Земли: палеобиогеографические аспекты. М.:Наука, 2005. 512 с.
- Ильин Р.С. Происхождение лессов. М.:Наука, 1978. 235 с.
- Керженцев А.С. Функциональная экология. М.:Наука, 2006. 259 с.
- Керженцев А.С. Новое перспективное научное направление. Вест. РАН, 2012, т.82, №5, с. 432-440.
- Ковда В.А. Биосфера и человечество. с.7-52. (Доклад ЮНЕСКО. Париж,1968). Биосфера и ее ресурсы. М.:Наука, 1971. 312 с.
- Ковда В.А., Бугровский В.В., Керженцев А.С., Зеленская Н.Н. Модель трансформации органического вещества в почве для количественного изучения функции почвы в экосистемах. Докл. АН СССР, 1990, т.312, №3. с. 759-762.
- Коссович П.С. Основы учения о почве. Ч.11, вып.1. СПб, 1911.
- Кузнецов В.Г. Литология микробиолитов. Вестник РАН, 2015. Т.85, №12. с. 1092-1102.
- Личков Б.Л. Современный литогенезис на материковых равнинах. Изв. АН СССР, 1945. Сер. Географич. И геофизич., Т.1Х, № 5-6. с.547-564.
- Марчук Г.И., Кондратьев К.Я. Приоритеты глобальной экологии. М.: Наука, 1992. 278 с.
- Неуструев С.С. Почвенная теория лессообразования. Природа, № 1-3, 1925.
- Путеводитель научных полевых экскурсий. Под ред. Ю.Г.Чендева. М.-Белгород. БелГУ, 2016. 122 с.
- Соколов И.А., Таргульян В.О. Взаимодействие почвы и среды: почва-память и почва-момент. Изучение и освоение природной среды. М.:Наука, 1976. с. 150-164.
- Соколовский А.Н. Грунтознавство. Харьков-Днепропетровск, 1933.
Приложение

Фото 1.

Фото 2.

Фото 3.
|





